This post was written by Jessica Sheu, a recent graduate from the computer science Masters program at San Jose State University. Jessica has been developing tools to apply machine learning techniques to automatically classify and ID images of sinking particles.
Jessica presenting her beautiful data
This June, I attended my first Ocean Carbon and Biogeochemistry (OCB) workshop! I presented a 2-minute lightning talk to advertise my poster on the research Dr. Durkin and I have been working on since Summer 2018: image classification of marine particles. I’m glad I did, because after hearing my talk, people who felt that my research could be specifically useful to them, sought out my poster. If you will be presenting a poster at a future OCB workshop, I highly recommend presenting a lightning talk too.
My first OCB experience was amazingly positive. Being from a Computer Science background with not a lot of technical oceanography knowledge, I was initially nervous. However, everyone I met was incredibly welcoming, and all the talks were interesting. I most enjoyed the plenary session on calcification and the carbon cycle organized by Andrea Fassbender and Jessica Cross, just because it made the most sense to me with my current limited ocean knowledge. I also loved how there was a panel after each plenary session that was very inviting for discussion. It was interesting to observe how everyone was so honest with their opinions of their research and future directions, while also being extremely open to hearing others’.
The entire workshop was not only extremely well-organized, but also thoughtfully so; I could tell that every event was placed and planned with intention. The organizers made sure to provide adequately long breaks between sessions so that attendees could stretch their legs, socialize, and refill on coffee. Delicious breakfast and lunch (and, on some days, hors d’oeuvres or dinner) were catered every day, and spacious transportation to and from the workshop was provided. I particularly appreciated how the agenda of each day was different. If there was a long day of talks on one day, the next day would intentionally provide more free time. In addition, I appreciated how accommodating OCB was towards students or early-career attendees. They organized special sessions for us, including a casual mixer and panel. Because of the mixer, I was able to meet a lot of people in a similar stage as me, as well as have some familiarity with faces for the rest of the week.
I’m very thankful for the OCB community and my experience at this workshop, and I’m glad that this was the first ocean conference I have attended. I felt so welcomed by everyone I met, and it was so interesting learning about other research during the talks and poster sessions. I would recommend this workshop to anyone, and I hope to attend more OCB workshops in the future!
Mandatory cannoli break with Jessica and MLML student Cindy Michaud before the conference.
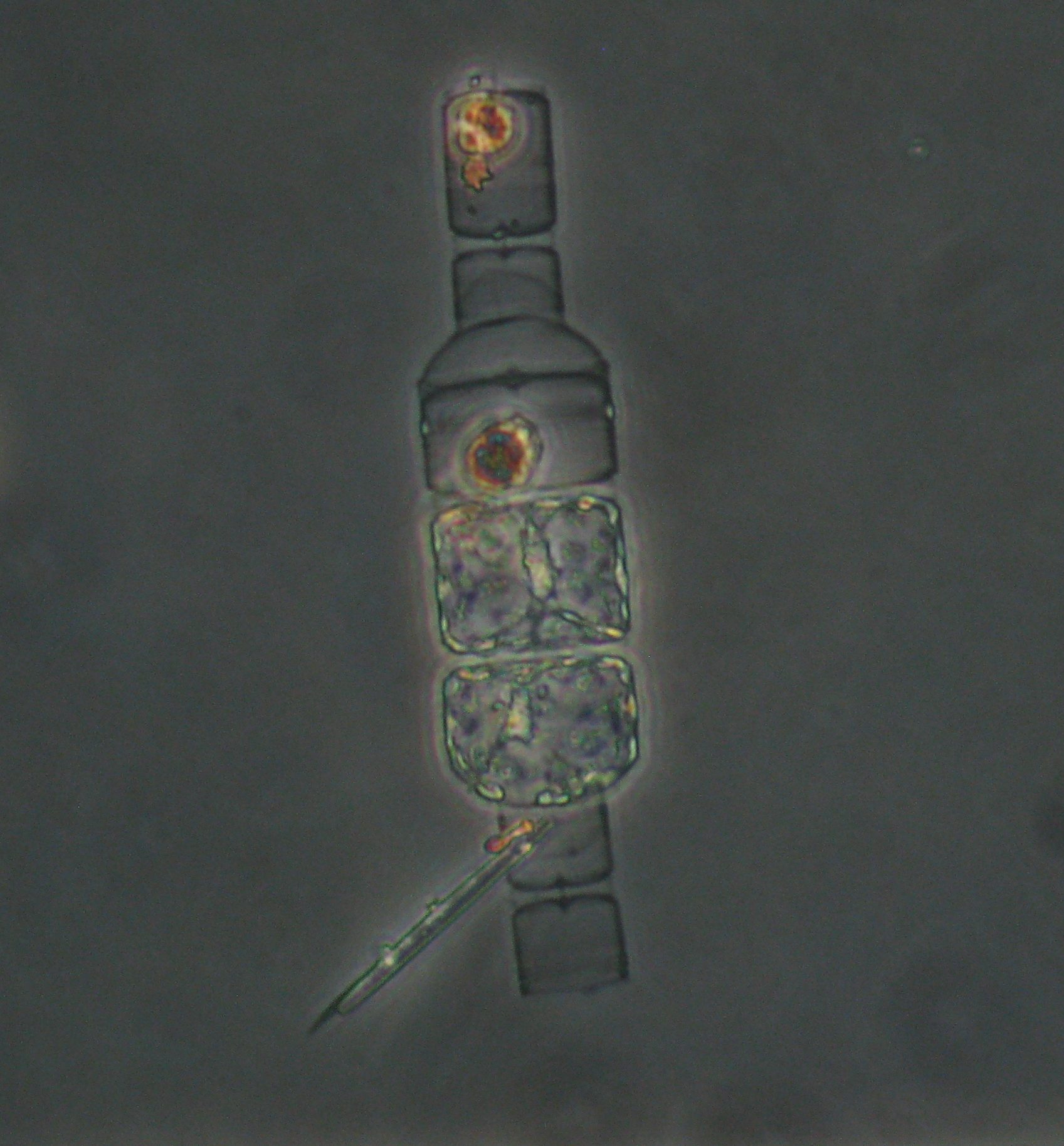
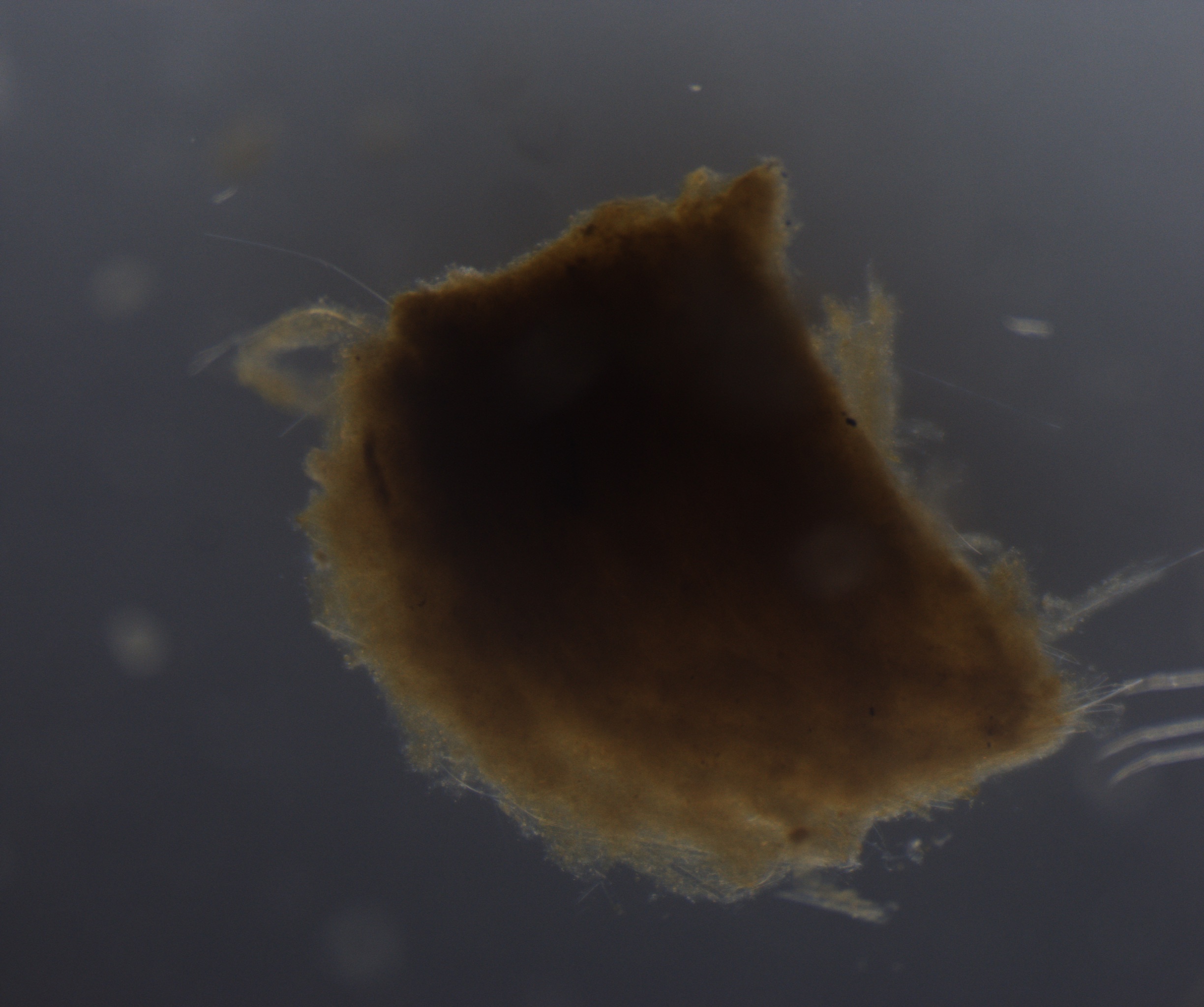
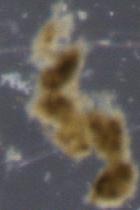
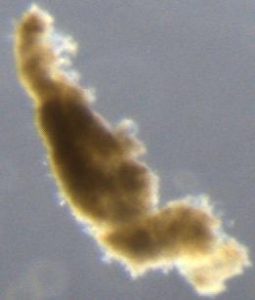
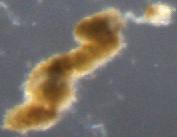
How do you tell the difference between “detrital aggregates” and “fecal pellets” (aka zooplankton poop)? In the last few months I have been looking through thousands of particle images, and categorizing them as fecal pellets or aggregates. These particles sink and transport carbon from the surface ocean into the “twilight zone” and sometimes all the way to the seafloor. We want to quantify how much of each particle type is sinking because they represents two very different ecological pathways of carbon sequestration in the ocean. Surprisingly, identifying these two particle types is not as straight forward as you might think. One reason is that some zooplankton make fecal pellets that look a lot like detrital aggregates. For example during the 2018 NASA EXPORTS research cruise, we encountered a larger population of vertically migrating salps. These gelatinous pelagic tunicates produce large fluffy pellets, pictured here:
Salp fecal pellet collected during 2018 NASA EXPORTSAnother salp fecal pellet. Notice the Rhizosolenia diatoms sticking out of the bottom right corner.
Fortunately salp pellets are fairly easy to spot since they are typically square shaped and look like shredded wheat cereal, according to Dr. Debbie Steinberg. Particles collected during the 2017 Sea to Space research cruise haven been a little more puzzling to me. These sinking particles collected offshore of the California coast look like fecal pellets, but also like more degraded aggregates (pictured below).
My current guess is that these are fecal pellets were produced by pyrosomes, another type of colonial pelagic tunicate. These bioluminescent animals lit up the ocean surface to a glowing green at night when our ship was steaming through them. Here is a micrograph of a colony of pyrosomes that got caught in the top of our sediment trap:
There are almost no studies on how pyrosomes digest their food, let alone what their fecal pellets look like. There is slightly more information on the closely related doliolids. I found a great video (linked to here) that shows how doliolids produce their fecal pellets. I recommend skipping ahead to minute 5. The video was created by Robert Fenaux, who also studied digestion in pyrosomes, but I can’t find that publication. These doliolid fecal pellets look very similar those particles I found in the sediment traps located where the pyrosomes were lighting up the water. As someone who considers herself a phytoplankton lover, I never thought I would get so excited about IDing poop particles!
Meg Estapa programing a trap in the Moss Landing Harbor
Last week I sailed on Moss Landing Marine Lab’s R/V John Martin for the first time! My collaborator Meg Estapa came to MLML to test neutrally-buoyant sediment traps that she has been building and programming in preparation for next month’s NASA EXPORTS research cruise. We wanted to perform a few more tests on these instruments before the big field campaign. Monterey Bay is a great location for this type of work because it only takes about 1 hour of steam time for a ship to get into very deep water (due to the underwater canyon, and the narrow continental shelf on the west coast). We need to be in deep water for this type of work because these instruments are designed to sink to a pre-programmed depth in the mesopelagic, drift with the currents for several days, and then resurface. Since we were testing how these instruments operate, we wanted to eliminate the risk of the instruments hitting the seafloor.
Cindy Michaud preparing to deploy a neutrally buoyant sediment trapJessica Sheu sailing out of Moss Landing harbor
Fortunately, we had help deploying and recovering these instruments from MLML students Cindy Michaud & Holly Chiswell, SJSU computer science student Jessica Sheu, and SJSU physics student Victoria Hickman. MLML’s Tom Connolly also represented from physical oceanography.
It does not escape my attention that we are testing these new sediment traps that measure carbon export on a ship named after John Martin. MLML’s John Martin and his students made some of the foundational observations and discoveries about the ocean’s carbon cycle and it is exciting to be continuing this type of research at MLML. Our work last week was successful thanks to the skilled captains of the R/V John Martin, John Douglas and Brian Ackerman.
For more on the action at sea and to learn more about the instruments, check out this video:
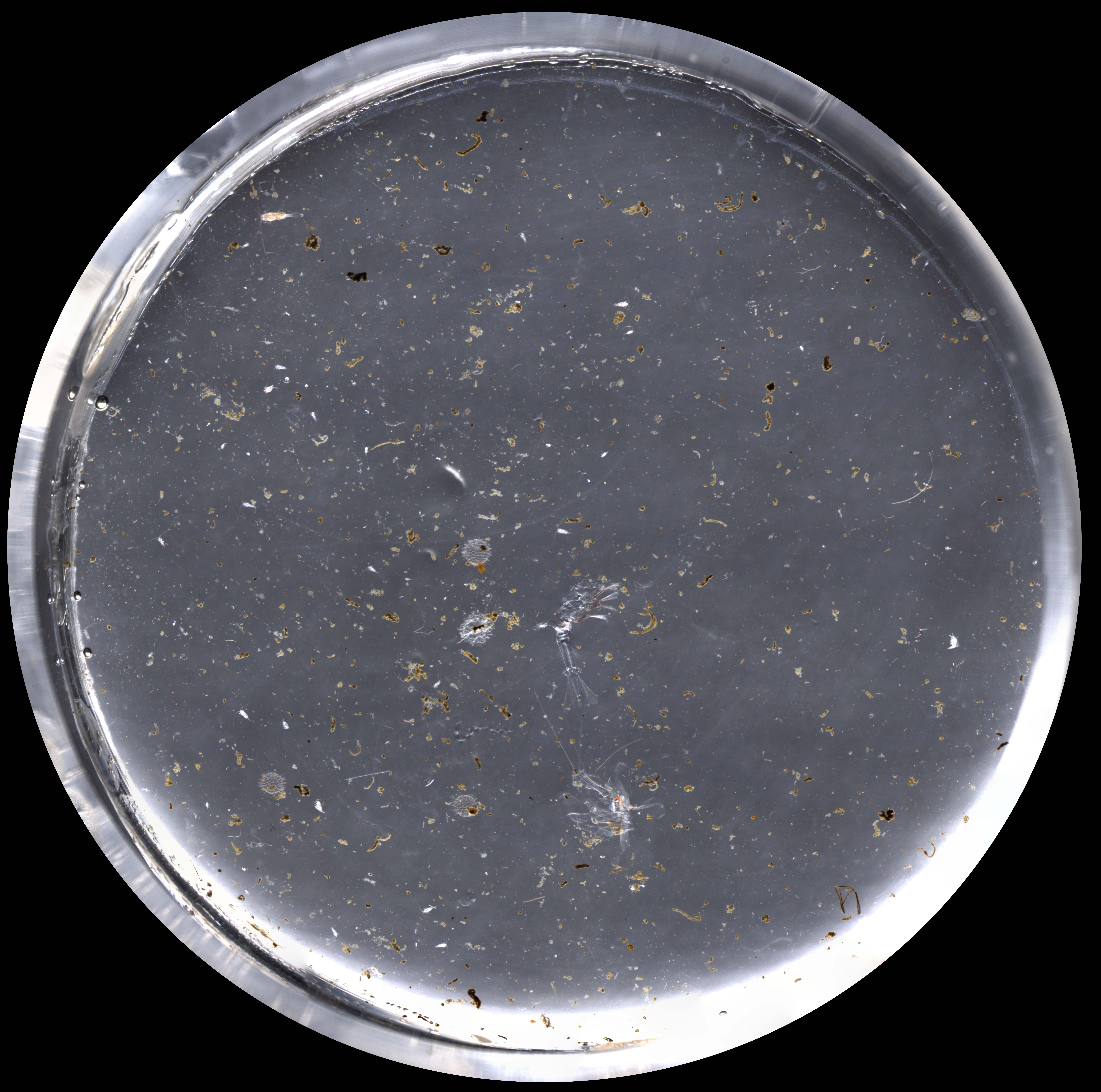
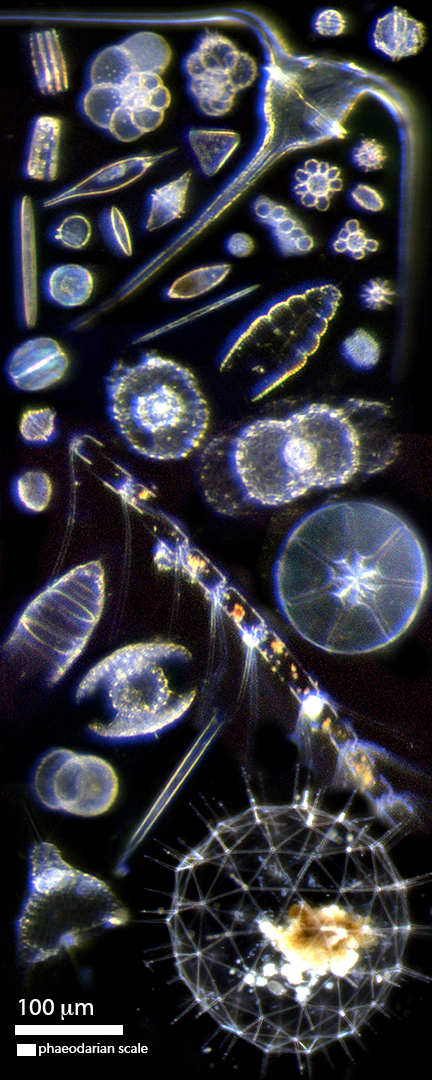
Last year I sailed on the Sea to Space Particle Investigation research cruise aboard the R/V Falkor. Our goal on that cruise was to connect surface phytoplankton communities with the amount of carbon exported out of the surface ocean and to identify the ecological mechanisms leading to their export. The most direct pathway between plankton and carbon export is direct settling of cells. On this cruise we deployed sediment traps containing polyacrylamide gel layers, like this one on the left. If you look closely, you will see individual particles (fecal pellets and aggregates) that sank into the gel. You can also see the remains of zooplankton that swam into the gel. When I examine the gel at higher magnification, I can see individual phytoplankton and protists (collage at the right). Because the gel isolates the particles as they fall into it, we can be relatively confident that these cells sank as individuals and not within larger detrital particles. Individual cells make up a small but consistent component of the total carbon export. On this cruise, they accounted for ~3-5% of total carbon flux at 150 m deep. The cells in the collage include dinoflagellates, coccolithophores, diatoms, foraminifera, radiolarians, and cilliates. The large spherical cell at the bottom is a phaeodarian, relative of the radiolarians, and is ~1 mm in size. Some of these protists are large enough to see in the gel image at low magnification.
Although most carbon is exported by detrital aggregations and fecal pellets, these individually sinking cells are fascinating. And beautiful.
At this year’s MLML open housee we set up some microscopes in the biological oceanography lab so that visitors could explore and discover plankton. In the morning before the open house I stopped at the wharf in Monterey to collect a phytoplankton net tow. It was a beautiful morning, and an otter with her pup swam by to check out what I was doing. Back at MLML we put the sample under a dissecting microscope and also projected live images on the wall at higher magnification from compound microscope. Most of the visitors had never seen plankton before, though many had heard of them. The comment I heard most often from the visitors was that they had no idea how small plankton were: most people expected them to be larger. Many scientists draw what they see while looking through the microscope. Drawing forces you to identify every detail of the cell, and helps you become better at identifying species-defining features. Plankton are also very beautiful, and inspire scientists to create art. I was inspired to share an art activity created by the scientific illustrator/artist Kirsten Carlson during a recent research cruise. Kirsten was the Schmidt Ocean Institute’s artist-at-sea during the R/V Falkor “Sea to Space” research cruise this past February (and a former MLML graduate student). She encouraged all the scientists draw plankton images on coasters while we were out at sea. For the open house I encouraged people (mostly kids) to illustrate what they were seeing under the microscope. People were very excited to attempt to translate what they were seeing onto paper. I was so fascinated by the particular details that stood out to different people. So people were focused on the way the light refracted off of the cell and affected the colors. Others focused primarily on shape. Some people were more interested in the big-picture view of how all the plankton looked mixed up together in the jar. Sharing how different people look at the same thing helps everyone discovery something new. This year we estimate ~2800 visited MLML for the open house. Here is a sample of what people saw under the microscope in the biological oceanography lab.
This week I received my lab’s first microscope! It is an Olympus SZX16 stereo microscope. This microscope will be used to image and quantify sinking particles collected in sediment traps. It also fits easily into a carry-on size bag so that I can safely transport it out to sea.
This microscope is coming with me on the upcoming Sea2Space cruise aboard the R/V Falkor (leaving next week!), and is part of a new research project recently funded by NSF. I received it just in time to test out its imaging capabilities with a preserved plankton net tow I collected last summer in Monterey Bay. This sample contained a type of gelatinous zooplankton called doliolids and a bloom of diatoms. I love how we can see both large and small things:
The doliolids are between 0.5 cm and 1 cm long, while the diatoms are about 0.005 cm long (or ~50 micrometers). The diatoms in this image are called Coscinodiscus.
I can’t wait to image sinking particles next month with my first microscope, and I’m already dreaming about microscope #2 for the lab.
A new project in my lab was recently funded by the National Science foundation. (My first NSF award!) The title of the project is:
“Collaborative Research: EAGER: Particle-specific DNA sequencing to directly observe ecological mechanisms of the biological pump”
This project is a collaboration with my co-principal investigators Margaret Estapa (Skidmore College) and Melissa Omand (University of Rhode Island) and will fund work during a month-long research cruise later this winter.
Like any good oceanographer, I immediately celebrated by purchasing a drill and a micropippetor.
Here is the non-technical abstract of the project: Carbon is fixed into organic matter by phytoplankton growing in the surface ocean, and is naturally sequestered in the ocean interior when particles and organisms sink: a process called the “biological pump”. Because of its recognized influence on the global carbon cycle, ocean scientists have studied the biological pump for decades. However, we still do not have a sufficient understanding of the underlying processes to accurately quantify and predict carbon cycling. Much of this uncertainty stems from an inability to directly link specific plankton in the surface ocean with the types of particles sinking out of the surface ocean. To address this missing link in biological pump research, this work will directly observe how plankton are transported out of the surface ocean using novel, particle-specific observational approaches embedded within an interdisciplinary field program that will finely resolve upper ocean plankton groups and the resulting amount of sinking carbon across space and in time. The genetic identity of organisms within different types of sinking particles will be determined by sequencing the genetic contents of individually collected particles. This new application of a molecular method will definitively link surface plankton with sinking particles at 5 locations across the Pacific Ocean. This work has the potential to transform our understanding of the biological pump by identifying previously unknown links between surface ecosystems and sinking carbon particles. Because this work is embedded within an interdisciplinary field program, including biogeochemical modelers and remote sensing scientists, these data will feed directly into new models of the biological pump, improving our ability to quantify and predict carbon uptake by the ocean.
This project was funded by the National Science Foundation Biological Oceanography program through their EAGER funding opportunity (Early-concept Grants for Exploratory Research).
We will be sailing on the Schmidt Ocean Institute’s vessel, the R/V Falkor, from Hawaii to Seattle. Along the way we will stop and deploy a variety of sediment traps and sensors to measure and collect sinking particles. This cruise is lead by chief scientist Ivona Cetinic (NASA) and will include an exciting interdisciplinary team of scientists including optical oceanographers, remote sensing scientists, and physical- chemical- and biological- oceanographers. The cruise has a website and is named the “Sea to Sky” cruise. If you want to check out the ship, of course the Schmidt Ocean Institute has a google-street-view-like virtual tour online. Here is the lab space we’ll be working in.
I am super excited about this project! Stay tuned for more updates as it progresses.
The evolution of silicon transporters in diatoms. Journal of Phycology(html)(pdf)
study co-authors: Julie Koester (UNC Wilmington), Sara Bender (Gordon and Betty Moore Foundation), E. Virginia Armbrust (UW)
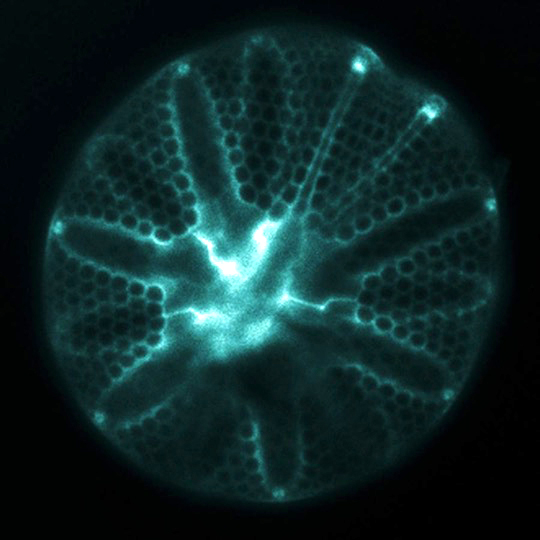
Fluorescently-labeled silica frustule of the diatom Asteromphalus
The ornately patterned silica cell walls (“frustules”) formed by diatoms attract attention for their beauty, but also give diatoms a distinct and globally important role in the oceans. These glass cell walls are created from dissolved silicon (silicic acid) that is actively taken up from seawater by the cells. Diatoms can not grow and divide without first forming this silica frustule, so their ability to take up silicic acid from seawater is a controlling factor on the magnitude of primary production that fuels ocean food webs and also influences global-scale biogeochemistry.
When diatoms evolved, the concentration of silicic acid in the ocean was likely much higher than today, and was drastically reduced as diatoms proliferated and diversified. The molecular machinery used to take up silicic acid may therefor include both relicts that evolved in an ancient ocean environment, and more recent adaptations that enable diatoms to thrive in the modern low-Si ocean.
We recently published a study examining the evolution of the proteins that enable diatoms to activley transport silicic acid into the cell: silicon transporters or “SITs”. This was made possible due to the recently sequenced, publicly available, community-produced transcriptomes of ~750 marine microeukaryotes. The diversity of organisms within this dataset enabled us to identify more diverse gene sequences across a broader range of organisms than previously possible.
First, we made a phylogenetic tree to model the evolutionary relationships among the gene sequences. In my opinion, this was the most challenging and critical part of the study because we wanted to use the most conservative methods to produce the most accurate tree, without sacrificing the inclusion of incredibly diverse sequences in the large transcriptome database. We eventually figured it out, but you’ll have to read the methods in the paper if you’re curious.
(Phylogeny of 473 SIT genes identified in this study)
This tree was much too big to analyze manually, which was a great motivation to learn python. (I used the ETE2 tools in python to study this tree.) Every “branch” on the tree is a gene sequence encoded by an organism, and branches that are connected are more closely related to one another. The nodes where branches connect represent a common ancestral gene that would have existed in the past.
One exciting discovery that emerged from this tree was the presence of “SITs” in other marine protists, not diatoms. These sequences clustered together, apart from the diatoms, but shared a common ancestral sequence with diatoms. This suggested that silicon plays a role in the cell biology of other marine protists (such as in a recent study of coccolithophores), and also that the earliest diatoms likely inherited SITs from an ancestral protist when they emerged in a high-Si ocean. Actively taking up Si is clearly an important function for the cell, even in high Si conditions, and we identified decendants of these earliest SIT genes in all major diatom lineages (black branches).
A second interesting discovery was that diatom SIT genes continued to diverge and diversify as diatom species evolved. When a gene duplicates in the genome, the duplicate sequence can change and has the potential to take on new functions. We identified 5 distinct types of SIT genes (branches on the tree are color-coded by type), and each species encodes its own repertoire of SIT gene types. The diatoms that evolved last (pennates) encode the most diverse combination of SIT sequence types. The diatoms that evolved first (centrics) encode the
least diverse combination of SIT sequence types.
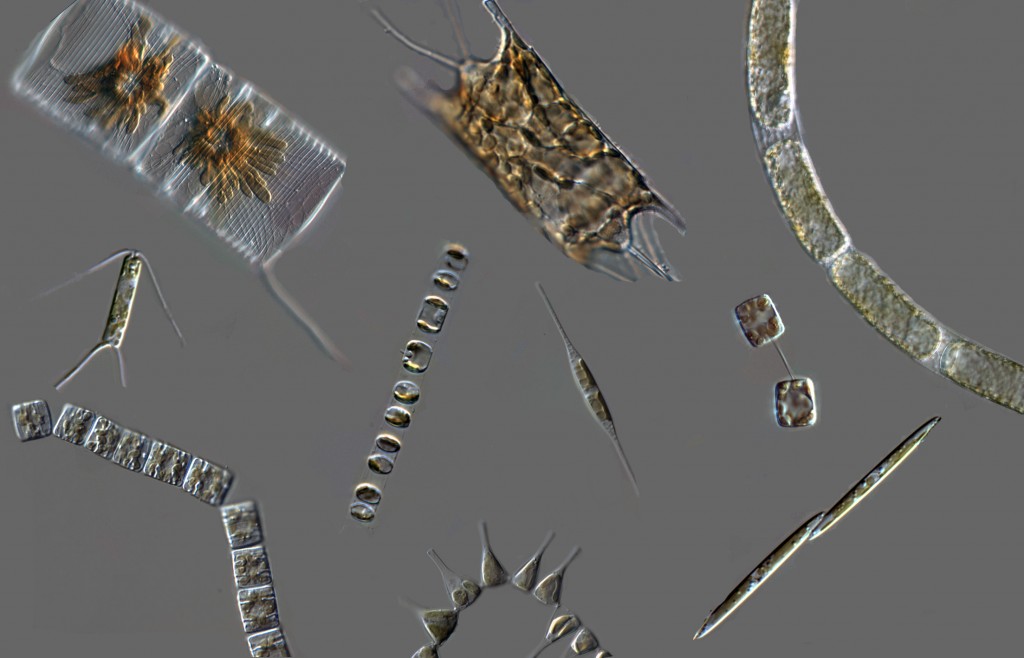
diverse diatoms
This is very exciting because of the potential ecological ramifications: different diatom lineages may have different abilities to take up silicic acid, and they may have distinct responses to changes in its availability. Encoding different SIT gene repertoires may be a molecular basis for niche-partitioning among co-exisiting species that need to utilize the same resource (Si).
The ecological implications of the evolutionary history of SITs were intriguing, and we wanted to know what type of SIT genes are used by natural communities of diatoms, and whether it changes depending on the species present and the environmental condition. To look for evidence that different SIT types are used in different conditions, we studied genetic samples from 4 locations off the coast of Washington and British Columbia (collected during the 2012 GeoMICS cruise). These samples represent all the genes transcribed (i.e.
“expressed”) by natural mixed plankton communities at the time of sampling (i.e. metatranscriptomes). Because we had such an extensive phylogenetic tree as a reference, we were able to identify environmental sequences by their SIT sequence type and also associate those sequences with organisms that have closely related sequences.
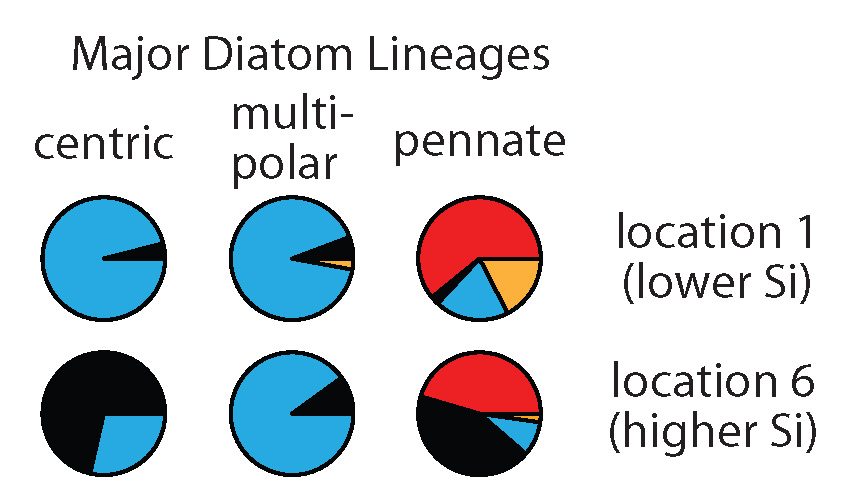
Pie charts show SIT sequence types (different colors) expressed by different diatom lineages at two different locations.
We found large differences in the type of SITs most expressed by different diatom lineages (compare pies horizontally). We also saw changes in the type of SITs expressed within these lineages between locations (compare pies vertically). Interestingly, at the location with the highest silicic acid, we detected relatively more of the earliest SIT type most closely related to the genes encoded by the first diatoms that lived in a high Si ocean (black pie pieces). In this study, we don’t have the data to definitively explain why diatoms changed the types of SITs expressed between environments, but the data do suggest that these measurements could be useful in future studies that try to resolve the responses and physiology of natural populations.
This project was a collaboration with the colleagues I worked with during my PhD at UW. We came up with the idea for this project just as we all moved to different parts of the country, but our enthusiasm for the topic is what kept us working on it over the next four years, often in our free time. Thank you to my “SIT-sters”!
Visiting Julie in Wilmington, NC to work on the SIT project. “SIT-in” T-shirts designed by S. Bender.
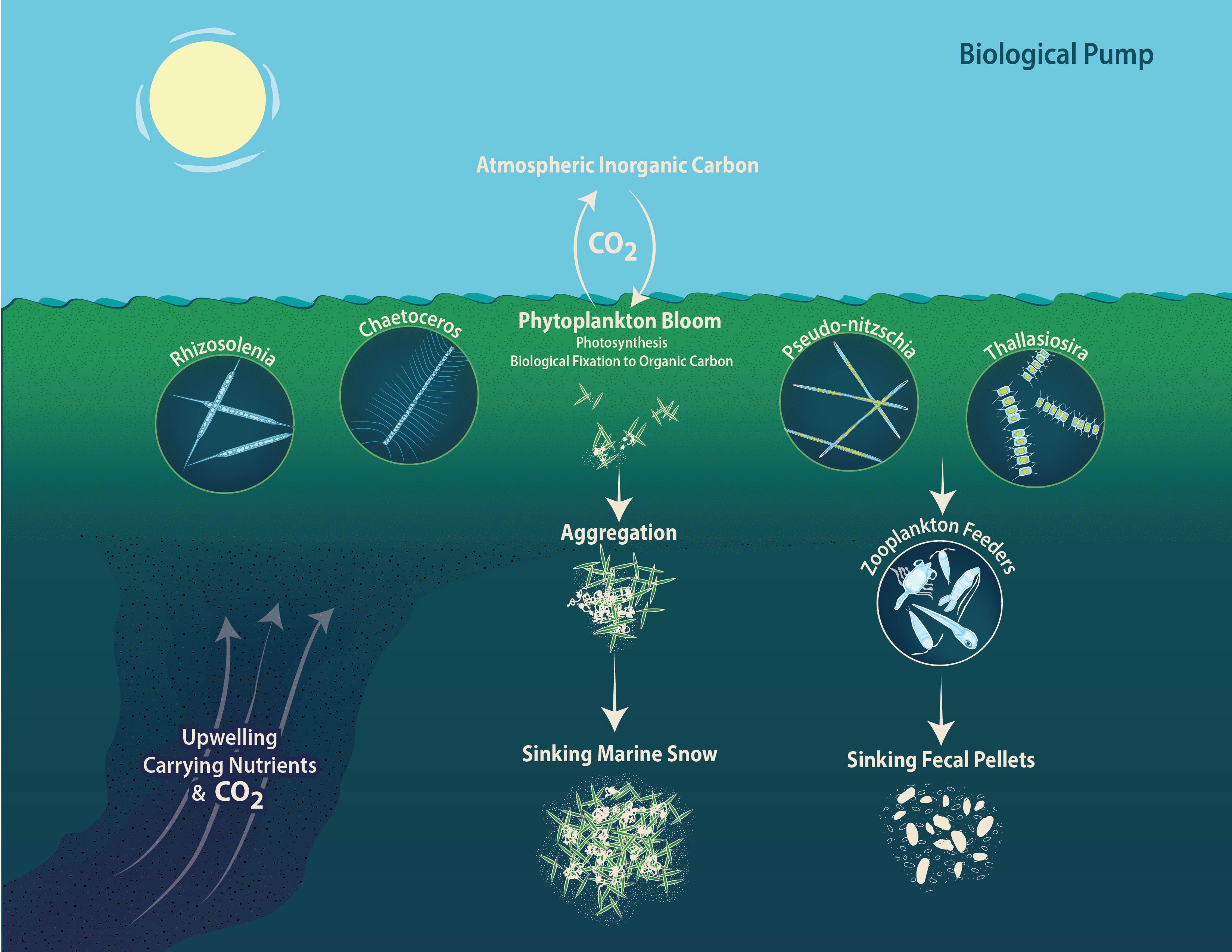
It is always exciting when science and art intersect. This summer, science illustrator Natalie Renier created this beautiful image to help CSUMB REU student Melia Paguirigan communicate her research about diatom aggregation:
The biological pump is very complex, and this image helped Melia communicate the components of that process that were directly related to her project.
To see more of Natalie Renier’s work, visit her website:
The main reason I enjoy working at the microscope is that I never know exactly what I will see. Today while counting a preserved phytoplankton sample collected at the Rhode Island shelf break I spotted these cells:
This is a chain of diatom cells (genus: Detonula, I think) undergoing a life cycle stage that allows them to get bigger. Diatom cells get smaller every time they divide. At some point, the cells are too small to divide anymore and must form either sexual stages or auxospores to get big again. In this photo the top cell in the chain is relatively small and does not appear to be vegetative (i.e. able to divide). The second cell down the chain has a small top half and a much larger bottom half. This cell formed an auxospore. The third and forth cells down the chain are large and contain cytoplasm and chlorophyll (the yellow pigment). These cells are probably the daughter (and grand-daughter) cells of that auxospore. Because the cells are in a chain, it is possible to see the history of this life cycle response.
I found a very similar image of Detonula auxospores here.





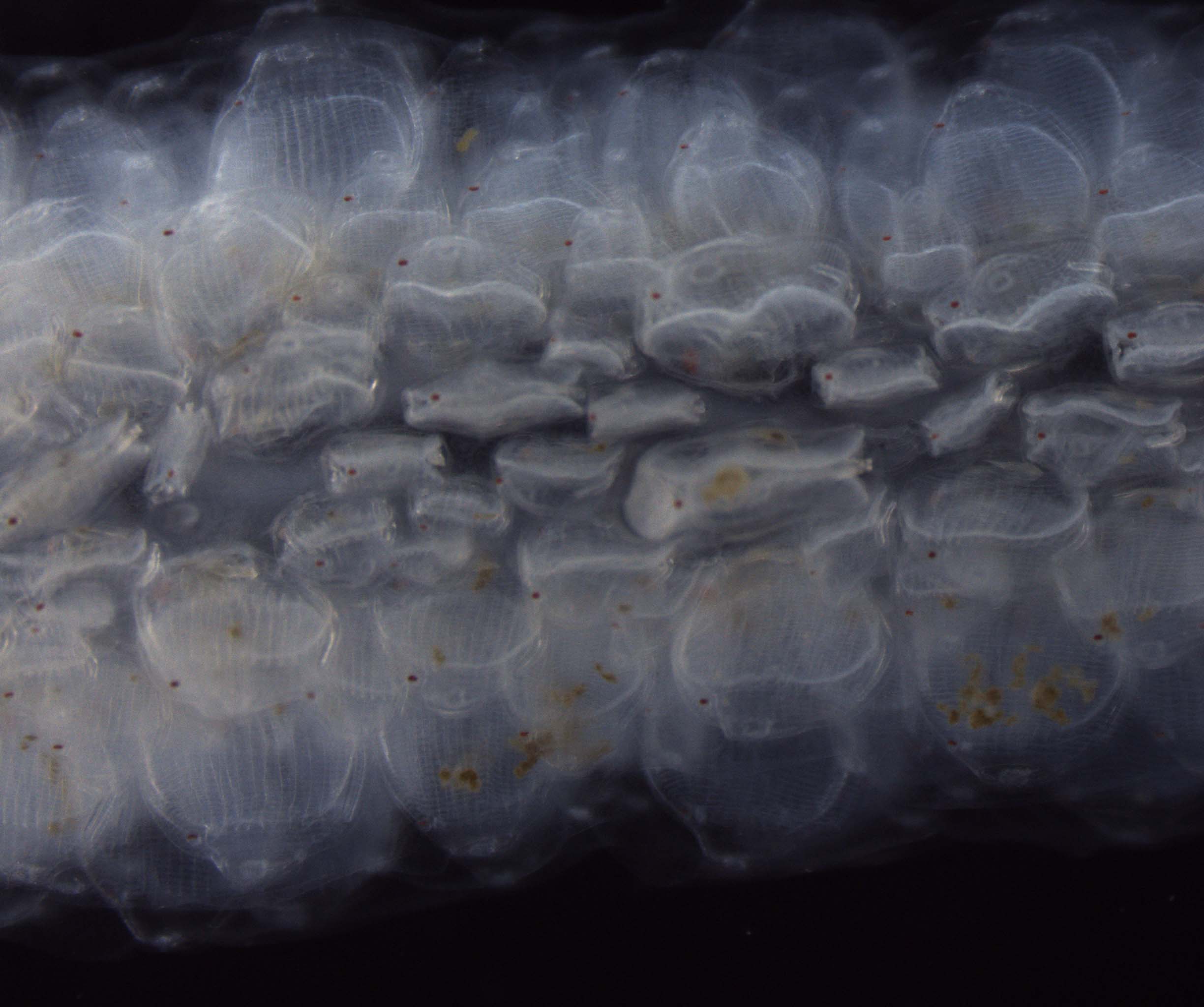



 At this year’s MLML open housee we set up some microscopes in the biological oceanography lab so that visitors could explore and discover plankton. In the morning before the open house I stopped at the wharf in Monterey to collect a phytoplankton net tow. It was a beautiful morning, and an otter with her pup swam by to check out what I was doing.
At this year’s MLML open housee we set up some microscopes in the biological oceanography lab so that visitors could explore and discover plankton. In the morning before the open house I stopped at the wharf in Monterey to collect a phytoplankton net tow. It was a beautiful morning, and an otter with her pup swam by to check out what I was doing.  Back at MLML we put the sample under a dissecting microscope and also projected live images on the wall at higher magnification from compound microscope. Most of the visitors had never seen plankton before, though many had heard of them. The comment I heard most often from the visitors was that they had no idea how small plankton were: most people expected them to be larger.
Back at MLML we put the sample under a dissecting microscope and also projected live images on the wall at higher magnification from compound microscope. Most of the visitors had never seen plankton before, though many had heard of them. The comment I heard most often from the visitors was that they had no idea how small plankton were: most people expected them to be larger.  Many scientists draw what they see while looking through the microscope. Drawing forces you to identify every detail of the cell, and helps you become better at identifying species-defining features. Plankton are also very beautiful, and inspire scientists to create art.
Many scientists draw what they see while looking through the microscope. Drawing forces you to identify every detail of the cell, and helps you become better at identifying species-defining features. Plankton are also very beautiful, and inspire scientists to create art.  I was inspired to share an art activity created by the scientific illustrator/artist
I was inspired to share an art activity created by the scientific illustrator/artist 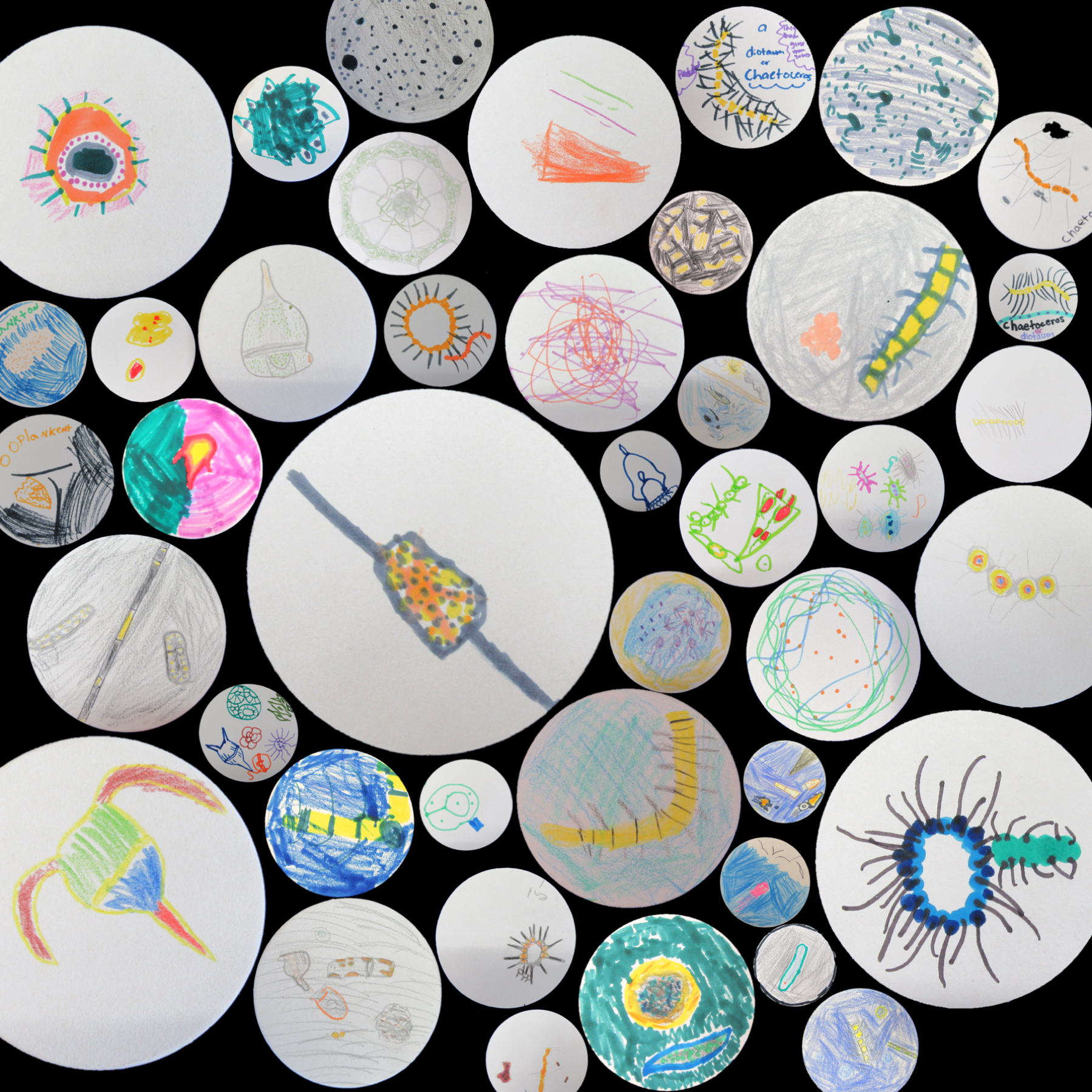
 This microscope will be used to image and quantify sinking particles collected in sediment traps. It also fits easily into a carry-on size bag so that I can safely transport it out to sea.
This microscope will be used to image and quantify sinking particles collected in sediment traps. It also fits easily into a carry-on size bag so that I can safely transport it out to sea.


 Like any good oceanographer, I immediately celebrated by purchasing a drill and a micropippetor.
Like any good oceanographer, I immediately celebrated by purchasing a drill and a micropippetor.